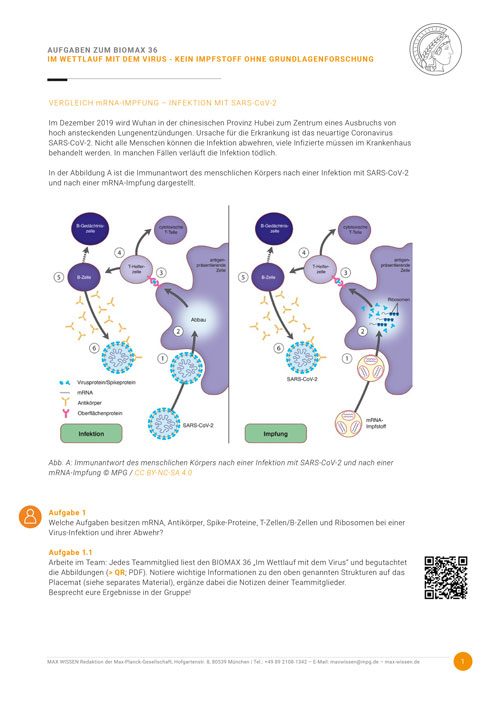
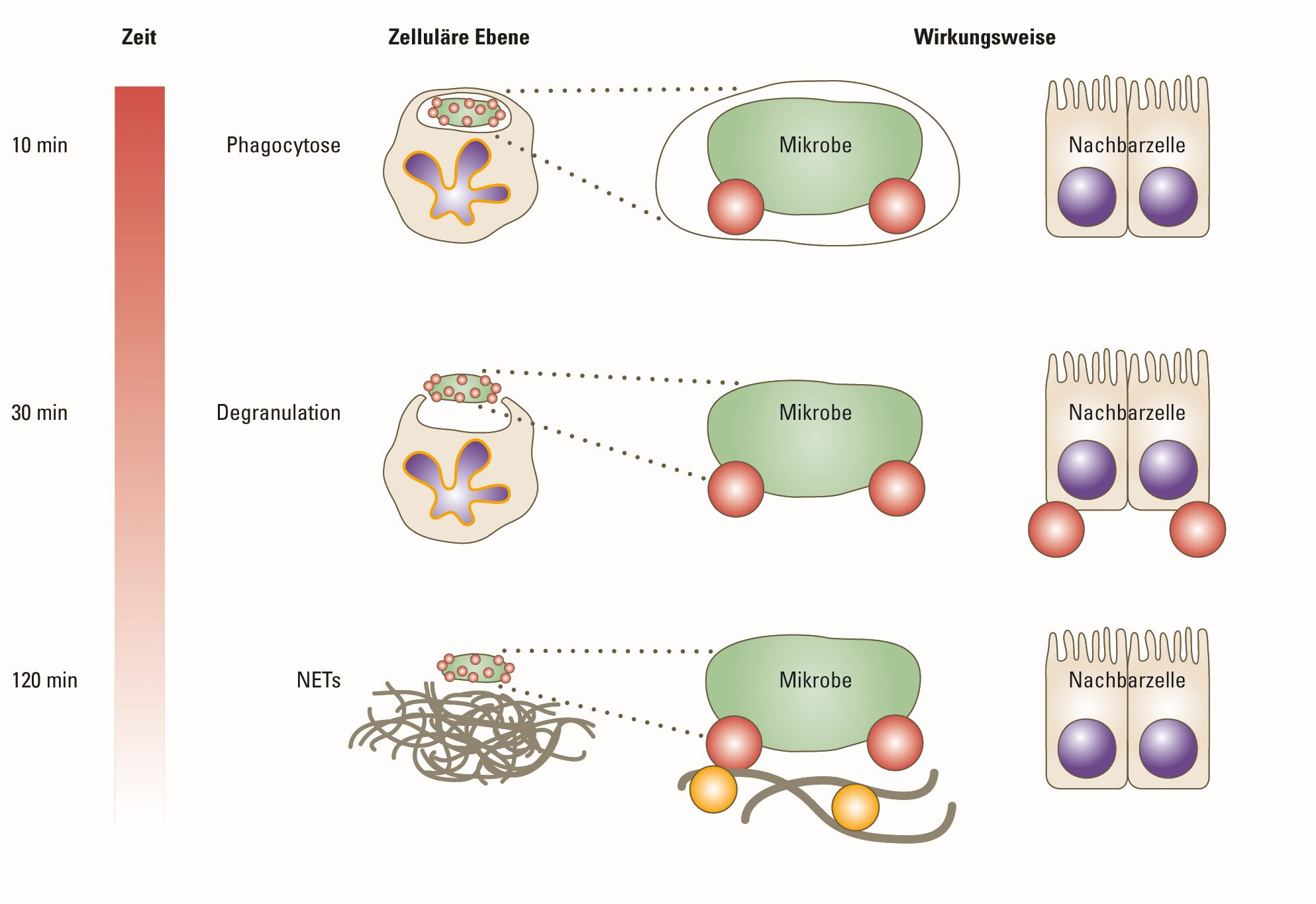
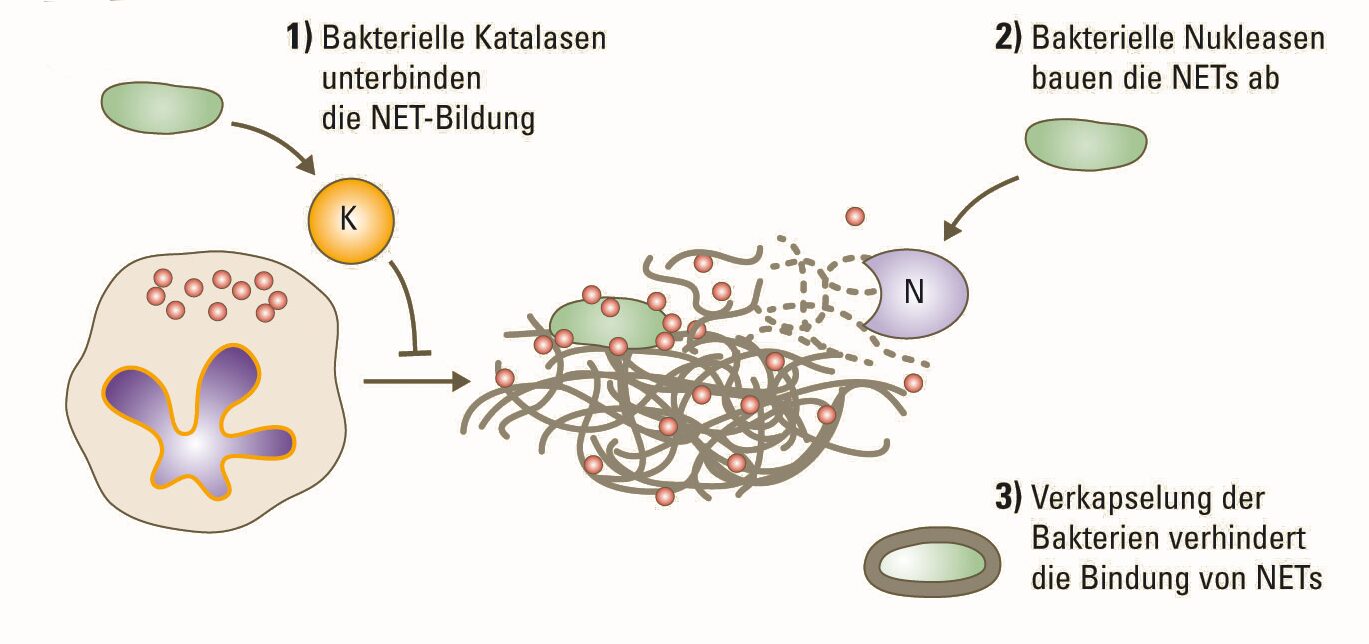
In den Körpern aller Lebewesen tobt ein täglicher Kampf gegen Viren, Bakterien, Pilze und Parasiten, die von außen eindringen. Selbst einfachste Organismen besitzen daher eine vielfältig ausgestaltete Immunabwehr. Die sogenannte angeborene oder natürliche Immunität, die sich schon bei wirbellosen Tieren und selbst bei Pflanzen findet, erlaubt eine besonders rasche, unspezifische, aber trotzdem wirkungsvolle Verteidigung. Die Struktur der beteiligten Proteine und die dazugehörige Bauanleitung sind im Genom festgelegt und können im Laufe des Lebens eines Individuums nicht an neue Bedingungen angepasst werden. Um diesen Nachteil zu beheben, hat sich im Verlauf der Evolution im angeborenen Immunsystem eine immer größere Zahl verschiedener […]